Anatomy & Physiology
Sea anemones, and all other cnidarians, are diploblastic animals. They have two germ layers, the endoderm and the ectoderm. The endoderm forms the gastrodermis and the ectoderm forms the epidermis. Sea anemones, and all cnidarian polyps, have a blind gut. Between the mouth and the coelenteron is the actinopharynx. Siphonoglyphs (ciliated groves) are located on the actinopharynx and create currents that bring oxygenated water into the coelenteron. Anemones can have one or two siphonoglyps, and are usually situated on the directive mesenteries. A. veratra has two siphonoglyphs. The actinopharynx leads into the coelenteron, which is continuous with the tentacles. The coelenteron (also referred to as the gastrovascular cavity) is the site of respiration, digestion and also acts as a hydrostatic skeleton. At the base of the animal is the pedal disc, which allows anemones to adhere to a substrate.
Musculature
The endodermal and ectodermal layers are largely made up of myo-epithelial cells, cuboidal epithelial cells with smooth muscle-like filaments at their base (Seipel & Schmid, 2006). These cells can have secretory, glandular or digestive functions (Seipel & Schmid, 2006). In addition to myo-epithelial cells, other types of muscle cells have been identified in cnidarians. These cells are typically flattened and elongated with basal myofilaments that may be smooth or striated, and can be arranged in two-dimensional sheets or three-dimensional bundles (Seipel & Schmid, 2006).
Anemones are simple animals and posses limited specialised muscular structures. Retractor muscles are orientated longitudinally and control column bending and shortening. Contraction of the oral disc is controlled by a spincter muscles, which are often located at the columns margin. Various muscular structures can be found on the mesenteries, and vary between species (Figures 1-2).
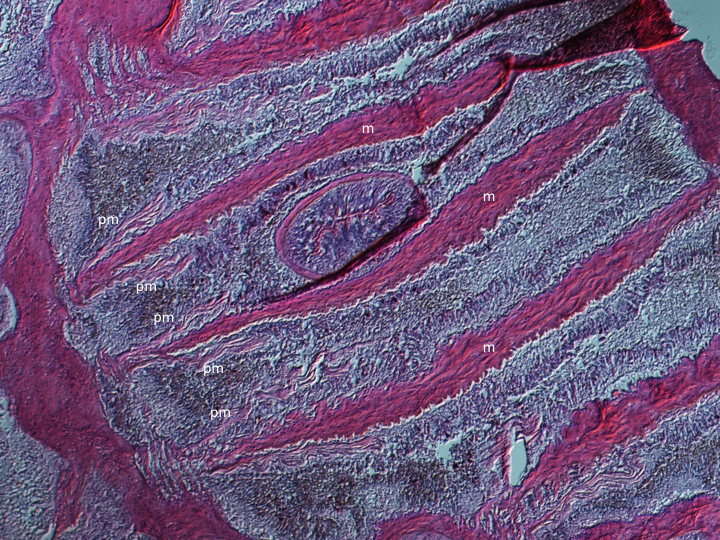
Figure 1: H&E stained cross section of an A. veratra column. m: mesentery, pm: parietal muscle.
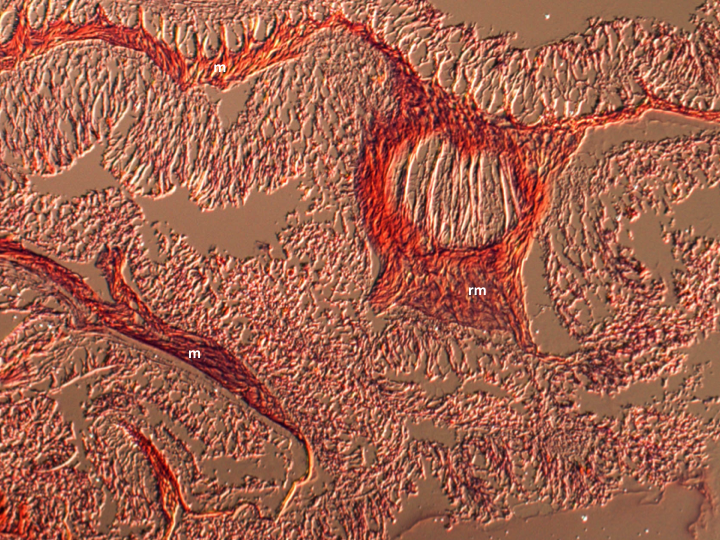
Figure 2: H&E stained cross section of an A. veratra column. m: mesentery, rm = retractor muscle.
Nervous system
Cnidarians posses diffuse, interconnected nerve nets at the base of the gastrodermis and the epidermis (Hickman et al., 2011). Unlike the nerve cells of most animals , which can only conduct signals in one direction, cnidarian nerve cells appear to be capable of bi-directional signalling (Hickman et al., 2011).
Mesenteries
Mesenteries are endodermal infoldings that extend from the body wall into the coelenteron (gastrovascular cavity)(Figure 3). The number, arrangement and type of mesenteries can be used, in part, to identify anthozoan species. Complete mesenteries extend from the column and are attached to the actinopharynx, incomplete mesenteries are not attached to the actinopharynx. Complete mesenteries can be directive or non-directive. Directive mesenteries are positioned on the directive axis. Mesenteries increase the surface area of the coelenteron and serve as points of attachment for muslces (longitudinal, transverse and sometimes basilar, depending on the species)(Figures 1-2) and gonads (Hickman et al., 2011; Moore, 2006).
In A. veratra mesenteries are hexamerously arranged. A maximum of five orders have been reported (Edmands & Fautin,1991). The first three orders are complete, while the higher orders are confined to the proximal end and may be rudimentary. (Edmands & Fautin, 1991). Mesenteries are added from the proximal end and are narrower distally (Edmands& Fautin, 1991). Gonads are present on all stronger mesenteries, except the directive mesenteries (Edmands & Fautin, 1991). A. veratra has two, symmetrical siphonoglyphs that are each connected to directive mesenteries (Edmands & Fautin, 1991).
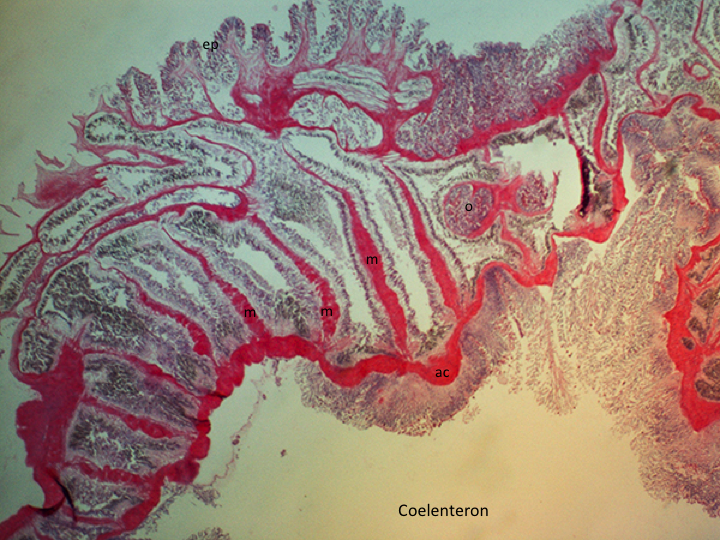
Figure 3: H&E stained cross section of an A. veratra column. m: mesentery, ac: actinopharynx, ep: epidermis, o: ovary.
Nematocytes
Nematocytes are a cell type, used in prey capture and defence, which defines the Cnidaria. A nematocyte contains a nematocyst, a subcellular organelle that is comprised of a capsule containing an attached hollow, coiled tubule. Nematocysts often contain toxins to kill or immobilise prey and predators.
Over 25 types of nematocytes have been described in the Cnidaria (Kass-Simon & Scappaticci, 2002). Nematocysts are single-use organelles. In order to conserve energy, often appropriate chemical and mechanical stimulation is required to trigger firing (Kass-Simon & Scappaticci, 2002). When nematocysts are fired, the everted tubule pierce, entwine or stick to their target (Kass-Simon & Scappaticci, 2002). Nematocyst discharge is one of the fastest movements in the animal kingdom and can be as short as 700 nanoseconds (Nüchter et al., 2006).
A. veratra has four types of nematocysts. Basotrichs are the most common and are found on tentacles, the actinopharynx, mesenterial filaments and the column. In addition to basotrichs, tentacles are armed with spirocysts, mesenterial filaments with microbasic p-mastigophores, and the column with holotrichs (Edmands and Fautin, 1991)(Table 1).
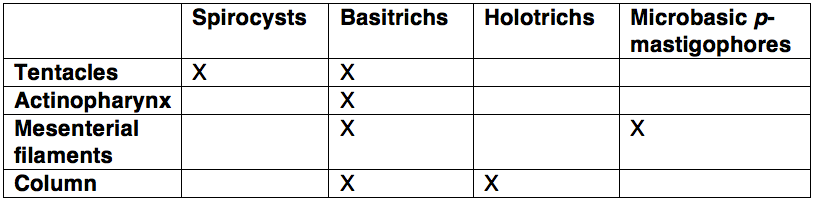
Table 1: Distribution of nematocyte types in A. veratra. Adapted from Edmand & Fautin (1991).
|